|




Een initiatief van :
Stichting Food-Info

|

|
Food-Info.net> Wetenswaar > Tomaat > Lycopeen
Lycopeen
Dit deel van Food-Info.net is gebaseerd op het verslag "Lycopeen, de gezonde kleur van tomatensap", gemaakt in het kader van het vak Geintegreerde Levensmiddelentechnologie (Wageningen Universiteit, 2002).

Aanwezigheid van lycopeen in de tomaat
Lycopeen wordt van nature gesynthetiseerd in de chloropasten van de tomaat. Deze groene chloroplasten veranderen in rode chromoplasten omdat het lycopeengehalte stijgt. Lycopeen zorgt voor 70 tot 80 procent van al het rode pigment in de tomaat. In rijpe tomaten komt lycopeen voor als lange, naaldachtige kristallen, waaruit grote delen van de chromoplasten bestaan (Nir et al. 1993). Deze kristallen ontstaan doordat de lineaire lycopeenmolekulen tegen elkaar aan gaan liggen en door onderlinge interactie een microkristallijn gebied vormen. Doordat het licht op deze gebieden reflecteert, krijgt de tomaat zijn typische rode kleur. Wanneer deze chromoplasten kapot worden gemaakt en lycopeen oplost in lipiden of andere 'oplosmiddelen', verandert de kleur van rood naar oranje.
Globulaire chromoplasten, die hoofdzakelijk beta-caroteen bevatten, zijn aanwezig in de gelei-achtige gedeelte van het pericarp (zie onderstaande figuur), terwijl de chromoplasten in het buitenste gedeelte van het pericarp omvangrijke kristallen van lycopeen bevatten (Nir et al. 1993).
Lycopeen komt overal in de tomaat voor (Ishida et al.,1998). McCollum (1955) heeft gevonden dat in het buitenste pericarp de grootste concentratie lycopeen voorkomt. In 1996 werd gevonden (Sharma en Le Maguer, 1996) dat de schil ook een rijke bron aan lycopeen (53.9 mg/100g) is en deze bevat ongeveer vijf keer meer lycopeen dan vermalen tomaten (11 mg/100g). Tomaten bestaan uit 5 tot 10 % droge stof, waarvan 75 % oplosbaar is. Van deze 5 tot 10% droge stof bestaat weer 1 tot 3% uit schil en zaden (Shi en Le Maguer, 2000). 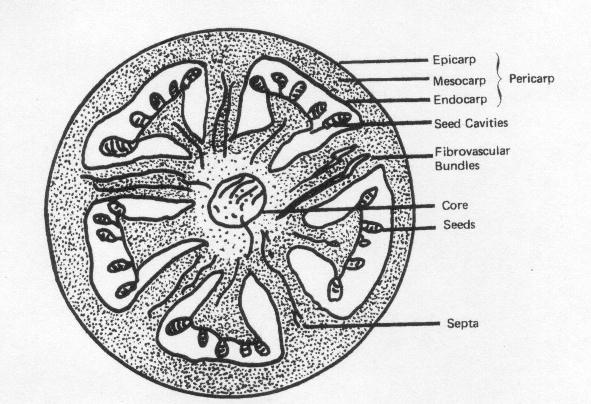
Doorsnede van een tomaat
Structuur, vorming en eigenschappen van lycopeen. Lycopeen, een terpenoid, wordt gevormd uit het C5-terpenoid isopentenyl difosfaat (IPP) (XVII) (Figuur 3). Dit molekuul wordt omgevormd tot geranylgeranyl difosfaat (C20) (XVIII). De dimerisatie van XVIII leidt tot phytoeen (7,8,11,12,7',8',11',12'-octahydro-gamma,gamma-caroteen) (XIX) en de stapgewijze dehydrogenatie via phytoflueen (15Z,7,8,11,12,7',8'-hexahydro-gamma,gamma-caroteen (XX), zetacaroteen (7,8,7',8'-tetrahydro-gamma,gamma-caroteen (XXI) en neurosproeen (7,8-dihydro-gamma,gamma-caroteen) (XXII) geeft lycopeen (I). 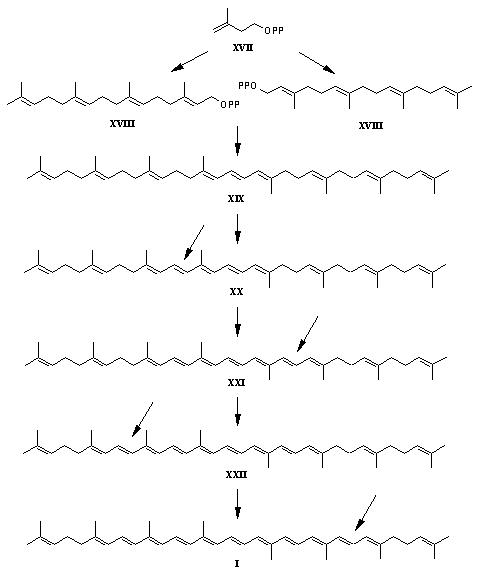
Figuur 3: Vorming van lycopeen in de tomaat Vanuit lycopeen worden overige carotenoïden, zoals beta-caroteen, gevormd door verschillende cyclisatiestappen. Met zijn acyclische structuur, het grote aantal geconjugeerde dubbele bindingen en zijn extreme hydrofobiciteit, bezit lycopeen veel unieke en aparte eigenschappen (Shi en Le Maguer, 2000). Cis/trans-isomerisatie
In de natuur komt lycopeen voor in de 'all-trans' vorm, welke thermodynamisch de meest stabiele vorm is. Door verschillende bereidingsstappen, zoals verhitten, kunnen enkele van deze transbindingen door isomerisatie overgaan in de mono- of polycis vorm. Deze cis-isomeren hebben andere eigenschappen dan hun trans vorm. Zo kunnen de cis-isomeren geen microkristallijne gebieden meer vormen, waardoor ze beter in vet oplosbaar zijn (Shi en Le Maguer, 2000). Tevens hebben de cis-isomeren een meer oranje-achtige kleur. De cis-isomeren onderscheiden zich ook nog van de 'all-trans' vormen in de volgende punten : lagere smeltpunten, kleinere extinctiecoëfficiënten, een verschuiving van het (-maximum en het verschijnen van een nieuw maximum in het ultraviolette spectrum, de zogenaamde cis-piek (Nguyen en Schwartz, 1999).
Niet alle dubbele bindingen kunnen overgaan in de cis-vorm wegens sterische hindering. Zo kunnen van de 11 geconjugeerde dubbele bindingen, er maar zes overgaan in de cis-vorm (Figuur 4). De 15-cis, 11-cis, 9-cis en de 5-cis-lycopeenvorm komen in het menselijke plasma voor, waarvan de 9-cis-vorm het meest aanwezig is (Anonymous, 1999). In onderzoek wordt meestal geen onderscheid gemaakt tussen de verschillende cis-vormen. Als er in het vervolg wordt gesproken over de cis-vorm van lycopeen, dan wordt niet één bepaalde vorm bedoeld. Isomerisatie van de trans naar de cis-vorm kan positief beïnvloed worden door verhitting, door dehydratatie en door verhitting uit te voeren in een vet medium in plaats van in water.
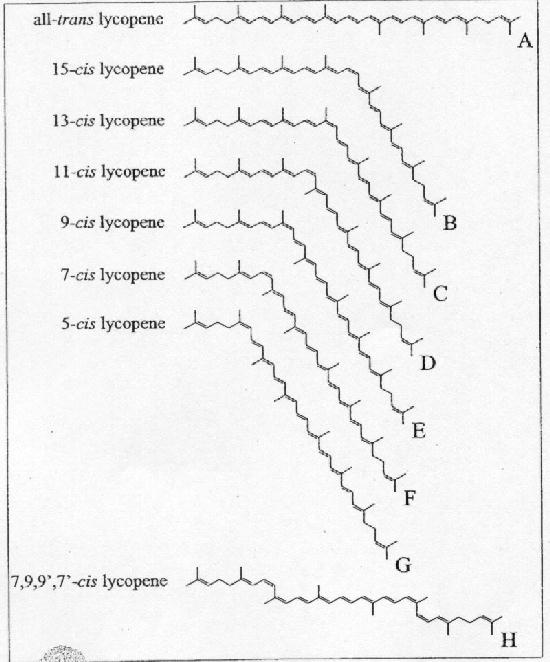
Figuur 4: De structuur van verschillende cis-isomeren van lycopeen (Nguyen en Schwartz, 1999) Reacties van lycopeen Reactief zuurstof kan DNA, eiwitten, koolhydraten en vetten beschadigen. De reacties van deze singlet zuurstof en peroxylradicalen zijn van belang omdat deze radicalen een belangrijke rol spelen in de oxidatie van onder andere vetten en ze dus indirect een rol speleln bij beschadiging van bijvoorbeeld membranen.
Carotenoïden staan bekend om hun anti-oxidatieve werking en lycopeen, als lipofiele antioxidant, bezit de hoogste snelheidsconstante kQ voor de reactie met singlet zuurstof in vergelijking met andere carotenoïden (Tabel 4). Hoe groter de snelheidsconstante kQ, hoe sneller de reactie verloopt (Di Mascio et al., 1989; Conn et al., 1991). Tevens heeft het aantal dubbele bindingen invloed op de snelheidsconstante. Lycopeen heeft meer dubbele bindingen dan ieder ander carotenoïde, inclusief beta-caroteen (Nir et al. 1993).
Tabel 4 : Vergelijking van antioxidant activiteiten van carotenoïden: Singlet zuurstof wegvanging, k Q (m -1 s -1) (Di Mascio et al ., 1989; Conn et al ., 1991)
Lycopeen |
|
Singlet zuurstof wegvanging, 10 9 *k Q |
31 |
Reactie van carotenoïdradicalen met O 2, 10 8 * k |
2 |
Singlet zuurstof wegvanging van andere carotenoiden, 10 9 * k Q |
|
gamma-Caroteen |
25 |
alpha-Caroteen |
19 |
beta-Caroteen |
14 |
Luteïne |
8 |
Astaxanthine |
24 |
Bixine |
14 |
Canthaxanthine |
21 |
Zeaxanthine |
10 |
De polyeenketen in het molecuul is een elektronenrijk en zeer reactief onderdeel, dat gevoelig is voor elektrofiele reagenten. Dit deel is met name verantwoordelijk voor de instabiliteit van de carotenoïden wat oxidatie betreft en is een belangrijk onderdeel van het molecuul als gekeken wordt naar de vrije radicaalreacties (Britton, 1995). Lycopeen vangt als antioxidant de singlet zuurstof en peroxylradicalen (ROO ·) weg volgens een tweede orde reactie. Wanneer lycopeen in aanraking komt met zuurstof of peroxylradicalen, kan het via verschillende mechanismen reageren. Een mogelijkheid is dat van het lycopeenmolecuul een H+ -atoom wordt afgesplitst, maar er vinden ook reacties plaats door middel van peroxylradicaaladditie of elektronenwegvanging (Woodall et al., 1997). Singlet zuurstof kan worden weggevangen door de volgende reactie, waarna het lycopeenradicaal via een onschadelijke reactie weer tot lycopeen wordt omgevormd :
LYC + O2-·
®
LYC·
+ O2 LYC·
®
LYC
In lycopeenradicalen is het ongepaarde elektron sterk gedelokaliseerd over de hele geconjugeerde keten van het molecuul. Dit heeft een stabiliserend effect en maakt verdere reacties, zoals bijvoorbeeld additiereacties, mogelijk op veel verschillende plaatsen aan het molecuul. De lycopeenradicalen kunnen worden gedetecteerd door hun specifieke spectrale eigenschappen, met een intense absorptie dicht bij het infrarood spectrum (Britton, 1995).
Peroxylradicalen zullen bijvoorbeeld worden weggevangen door de volgende reacties:
ROO·
+ LYC ®
ROO-LYC·
ROO-LYC·
+ ROO·
®
ROO-LYC-OOR, of: ROO·
+ LYC ®
ROOH + LYC·
Bij lage zuurstofconcentraties zal lycopeen vooral de peroxyradicalen wegvangen en zal het reageren als een verbreker van de propagatiereacties van radicalen. Bij hoge zuurstofconcentraties zal een lycopeenradicaal echter met zuurstof reageren en een carotenoïd-peroxy-radicaal vormen:
LYC·
+ O2 ®
LYC-OO·
Dit is een auto-oxidatieproces en het carotenoïd-peroxy-radicaal zou als pro-oxidant reageren, waardoor de peroxidatie van onverzadigd vet (VH) zal worden gestimuleerd en de schade dus zal worden vergroot (Britton, 1995):
LYC-OO·
+ VH ®
LYC-OOH + V·
V·
+ O2 ®
V-OO·
Carotenoïden inactiveren singlet zuurstof via 'fysische' (>99,9% van de reacties) of 'chemische' (<0,05%) wegvanging (Stahl en Sies, 1996). In het proces van de fysische radicaalreactie blijft het carotenoïd intact, zodat het nog meer radicaalreacties kan ondergaan. Bij het chemische wegvangen wordt lycopeen afgebroken tot andere verbindingen, waarbij ook de karakteristieke rode kleur van het molecuul zal verdwijnen (Sies en Stahl, 1998). Dit proces wordt 'bleken' genoemd. Intensievere lichtinval en temperatuur staan direct in correlatie met lycopeendegradatie in de aanwezigheid van zuurstof (Nguyen en Schwarz, 1999). Bepaalde metaalionen, zoals Fe3+ en Cu2+ katalyseren de afbraak van lycopeen.
Er is maar weinig verschil in de kQ gevonden voor de 'all-trans', de 9-cis en de 15-cis-isomeren van beta-caroteen gevonden (Conn et al. 1991). Er wordt aangenomen dat dit ook zal gelden voor de kQ van de verschillende lycopeenisomeren.
Een andere functie van carotenoïden is dat ze cellen beschermen tegen foto-oxidatie door te reageren met de aangeslagen fotosensitisers. Deze aangeslagen fotosensitisers kunnen ook weer singlet zuurstof vormen, dat weer oxidatiereacties op gang brengt (Woodall et al.1997). In het bloedplasma hebben lycopeen en (-caroteen een grotere weerstand tegen foto-oxidatie dan luteïne of zeaxanthine (Stahl en Sies, 1996).
Doordat lycopeen een lineaire structuur heeft en geen beta-ionenringen bevat, bezit het geen pro-vitamine A activiteit.
Invloed van ras op lycopeengehalte Van tomaten zijn rode en gele rassen bekend. In de gele rassen wordt de kleur voornamelijk veroorzaakt door beta-caroteen, in de rode varianten is lycopeen de veroorzaker van de kleur. Om lycopeen als natuurlijke kleurstof te gebruiken voor commerciële toepassingen, worden extra rode tomaten gekweekt, die een hoger lycopeengehalte hebben dan de meeste industriële soorten (Tabel 5) (Scott en Hart, 1995; Shi en Le Maguer, 2000)
Door klassieke veredelingstechnieken kunnen nieuwe hybride tomaten soorten ontwikkeld worden, die drie tot vier keer zoveel lycopeen bevatten als de standaard tomaat.
Tabel 5: Lycopeen- en b-caroteengehaltes in 12 tomatensoorten (mg/100 g) (Scott en Hart, 1995)
|
Totaal Lycopeen |
Trans Lycopeen |
Trans beta-caroteen |
Rode varianten: |
|
|
%1 |
|
Cherry |
3,8 |
2,7 |
(71) |
0,5 |
'Large' |
2,3 |
1,9 |
(84) |
0,3 |
'Salad' |
2,5 |
2,2 |
(85) |
0,5 |
Flavourtop |
5,7 |
5,0 |
(88) |
0,4 |
Tigerella |
1,6 |
1,2 |
(77) |
1,7 |
Ida F1 hybrid |
1,7 |
1,3 |
(77) |
1,0 |
Shirley F1 |
2,3 |
2,1 |
(89) |
0,8 |
Craig |
3,9 |
2,9 |
(75) |
1,1 |
Moneymaker |
4,3 |
3,5 |
(82) |
0,4 |
Allicanti |
4,0 |
3,7 |
(91) |
0,5 |
Beefsteak |
4,8 |
2,7 |
(56) |
0,9 |
Gele variant: |
|
|
|
|
Sungold |
0,5 |
0,4 |
(74) |
2,2 |
Gold Sunrise |
0,02 |
0,02 |
(100) |
0,09 |
1 : gedeelte translycopeen als percentage van het totaalgehalte aan lycopeen
Er bestaan ook nog enkele rassen, die nog aanzienlijk meer lycopeen bevatten (VT-145-7879: 30 mg/100g; Fresh tomato pear type (Solanum lycopersicum, Mill): 54-70 mg/100g), dan de rassen die in tabel 5 staan.Het nadeel van deze lycopeenrijke rassen, is dat deze nog niet zoveel opbrengst leveren per hectare, waardoor het nog erg duur is om deze voor commerciële doeleinden te gebruiken.
Invloed van rijping op het lycopeengehalte Het gehalte aan antioxidanten in tomaten hangt af van zowel genetische- als omgevingsfactoren, als van de manier van rijping (Giovanelli et al., 1999). Het lycopeengehalte stijgt voornamelijk in de allerlaatste fase van de rijping van de tomaat (Davies, 1998). Heinonen et al. (1989) rapporteerden dat de lycopeenconcentratie in tomaten in de zomer (juni tot augustus) hoger is dan in de winter (oktober tot maart), respectievelijk 3,6-6,6 mg/100g en 2,6-3,1 mg/100g. Tomaten die in de kas groeien (ongeacht in de zomer of winter) bevatten een lager gehalte aan lycopeen dan tomaten die gedurende de zomer buiten groeien. Tomaten die bij 16 ºC buiten groeien hebben een lycopeen gehalte van 5,8 mg/100g, als diezelfde tomaten in een kas groeien is het lycopeen gehalte 2,0 mg/100g. Misschien is dit te verklaren doordat de lichtintensiteit in een kas niet zo groot is als die van zonlicht in de zomer, of het ligt aan het temperatuursverschil tussen binnen en buiten de kas.
Het rijpingsproces van tomaten kan op twee verschillende manieren plaatsvinden, namelijk rijping van de tomaat voor de oogst en rijping nadat de tomaat al geoogst is. In het eerste geval blijft de tomaat aan de plant totdat deze rijp en rood is; in het tweede geval wordt de tomaat nog groen geplukt, waarna deze in een goedgeventileerde ruimte van 20 °C met een licht- en donkercyclus worden bewaard (Giovanelli et al.,1999). Om de verschillen tussen de twee verschillend gerijpte tomaten te kunnen weergeven, kan de a*/ b* -waarde als referentiewaarde worden gebruikt. De a*/ b* -waarde komt voort uit de L*, a*, b*, -kleurbepaling en geeft de verhouding tussen de rode en groene component van de kleur weer en representeert de kleurindex, welke de kleurvariatie gedurende het rijpingsproces beter weergeeft. Yeatman (1969) gaf aan dat de a*/ b* -waarde een hoge mate van lineaire correlatie met visuele kleurscores van tomatenproducten vertoont. Als het lycopeengehalte tegen de a*/ b* -waarde wordt uitgezet in een grafiek, is er te zien dat de grafiek (Figuur 5) van de aan de plant gerijpte tomaat een lineair verloop heeft, terwijl de grafiek van de tomaat die na de oogst is gerijpt een exponentieel verloop heeft. Bij een a*/ b* -waarde van 2,5 is het lycopeengehalte van de laatstgenoemde zelfs twee keer zo veel. Hieruit valt te concluderen dat er tussen de op verschillende manieren gerijpte tomaten een verschil is in de lycopeenvorming en dat er meer lycopeen wordt gevormd in de tomaten die na de oogst zijn gerijpt. Uit deze gegevens komt ook naar voren dat bij eenzelfde a*/ b* -waarde van de tomaten, geen dezelfde lycopeengehalten hoeven te bestaan. De rode kleur van tomaten kan ook voor een deel ontstaan door b-caroteen, de kleurstof die na lycopeen het meeste voorkomt in de tomaat. 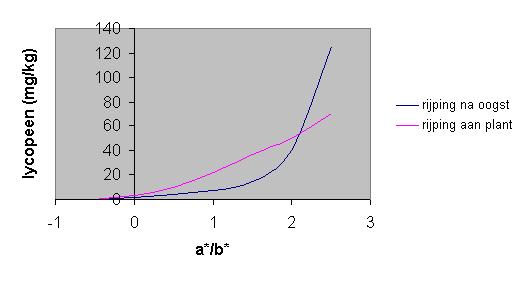
Figuur 5: Verschillen in lycopeengehalte ten opzichte van a*/b* in tomaten die aan de plant rijpen en in tomaten die na de oogst rijpen (Giovanelli, 1999)
Als de tomaten aan één en dezelfde plant rijpen, zal de mate van rijping tussen de tomaten verschillen door de positie aan de plant en daardoor door de blootstelling aan licht. De rijping na de oogst heeft ook als voordeel dat alle tomaten evenveel licht krijgen, waardoor de tomaten ongeveer tegelijk rijp zijn en dus een betere opbrengst opleveren. Er zijn ook nog onderzoeken uitgevoerd naar de invloed van bepaalde toevoegingen tijdens de rijping.
Ethyleen heeft wel invloed op de rijping en op de a*/ b*-waarden, maar niet op het lycopeengehalte (McDonald et al.,1996). Zoals hierboven beschreven heeft beta-caroteen ook invloed op de a*/ b*-waarde.
Er zijn verschillende onderzoeken in vitro uitgevoerd, maar deze zijn nog niet in vivo uitgevoerd en het is nog niet zeker of de stoffen mogen worden toegevoegd. Er zijn studies uitgevoerd met suspensieculturen waar 2-(4-chlorophenylthio)-triethylamine (CPTA) aan is toegevoegd (Robertson et al., 1995). Uit deze onderzoeken is gebleken dat de lycopeenconcentratie in verband staat met het toevoegen van CPTA en wel zo dat het lycopeengehalte stijgt. Boven een bepaalde concentratie (50-75 mg/l) CPTA stijgt de lycopeenconcentratie niet meer. Dit suggereert dat CPTA een alternatieve vorming van lycopeen mogelijk maakt. Tegelijkertijd werd gevonden dat er minder beta-caroteen wordt gevormd als er CPTA aan het medium is toegevoegd. Er is bekend dat oniumcomponenten, zoals CPTA, de cyclisatiereacties om (-caroteen te vormen uit lycopeen remmen (Ishida et al.,1998).
De rijping van een tomaat en hiermee ook de lycopeenconcentratie, wordt ook beïnvloed door de expressie van een AGAMOUS (TAG1) gen (Ishida et al., 1998). De TAG1-RNA-concentratie wordt verhoogd gedurende de rijping. Dit gen lijkt een optimale expressietemperatuur te hebben tussen de 16 en 23 °C, waaruit volgt dat lycopeenaccumulatie vooral plaatsvindt als de rijpingstemperatuur tussen deze waarden ligt (Ishida en Bieche, 1999).
Opslag tijdens rijping
Tomaten die na de oogst rijpen, worden snel overrijp en gaan achteruit in kwaliteit en houdbaarheid door opslag bij buitentemperatuur. Ook gaat veel lycopeen verloren na het oogsten (Türk et al,1994). De houdbaarheid kan verlengd worden door opslag bij lage temperatuur, maar een groot nadeel is dat bij lage temperatuur de tomaten niet volledig zullen rijpen. De tomaten worden niet rood en er wordt nauwelijks lycopeen gevormd. De opslagcondities moeten optimaal zijn, zodat de kwaliteit zo hoog mogelijk wordt en het bederf en het verlies aan lycopeen zo minimaal mogelijk zal zijn. Kwaliteit wordt hier bepaald door factoren als kleur en consistentie, maar het lycopeengehalte wordt hier niet tot de kwaliteitsfactoren gerekend. Het gehalte aan lycopeen en de kwaliteit wordt tijdens de opslag vooral beïnvloed door de combinatie van tijd en de omgevingstemperatuur.
Bij hoge omgevingstemperatuur wordt sneller lycopeen gevormd dan bij lage temperatuur. Als je groene tomaten, die nog moeten rijpen tijdens opslag, opslaat bij een temperatuur van 0 ºC en deze vergelijkt met tomaten die bij een temperatuur van 18 ºC worden opgeslagen, blijkt dat bij 18 ºC het lycopeengehalte hoger wordt. Opslag van drie weken bij 18 ºC geeft een betere kwaliteit dan opslag van drie weken bij 0 ºC. Zelfs als de tomaten zes weken worden opgeslagen bij 0 ºC hebben ze nog een slechtere kwaliteit en lager lycopeengehalte dan de tomaten die maar drie weken bij 18 ºC zijn opgeslagen. Bij een opslagtemperatuur van 18 ºC kunnen de tomaten maar drie weken bewaard blijven, omdat ze na deze tijd te rijp worden en tekenen van rotting gaan vertonen (Türk et al., 1994).
Bovenstaande geeft aan dat om een lange opslagtijd mogelijk te maken, de tomaten koel bewaard zouden moeten worden, maar dit heeft tot gevolg dat de tomaten niet voldoende rijpen. Een methode om wel een langere opslagtijd te creëren, maar toch de tomaat volledig te laten rijpen, is door de tomaten voor de opslag een hittebehandeling te laten ondergaan. Hitte behandeling voor de opslag verhoogt de synthese van lycopeen (Lurie en Klein, 1991). Hakim et al.(1996) hebben onderzoek gedaan naar de effecten van een hittebehandeling voor opslag. Uit dit onderzoek is gebleken dat de tomaten met hittebehandeling een betere kwaliteit bezaten dan de tomaten die zonder hittebehandeling waren opgeslagen. Verder bleek dat hoe hoger de behandelingstemperatuur was (onderzocht tot 39 ºC, 72 h.), hoe beter de kwaliteit was. Tevens was bij 39 ºC de concentratie lycopeen het hoogst bij de voorverhitte tomaten (Hakim et al., 1996). Biobeschikbaarheid van lycopeen.
Biobeschikbaarheid wordt gedefinieerd als het percentage van een ingenomen nutriënt dat geabsorbeerd wordt en daardoor beschikbaar is voor gebruik in normale functies en metabolische processen in het lichaam (Shi en Le Maguer, 2000). De absorptie van lycopeen uit het voedsel is erg wisselend en kan worden beïnvloed door verschillende factoren. Deze factoren zijn onder andere de verschillende chemische vormen van lycopeen (namelijk cis of trans), de dosis, factoren in het voedsel, fysiologische factoren, demografische factoren en gedragsfactoren (Johnson, 1998; Shi en Le Maguer, 2000)
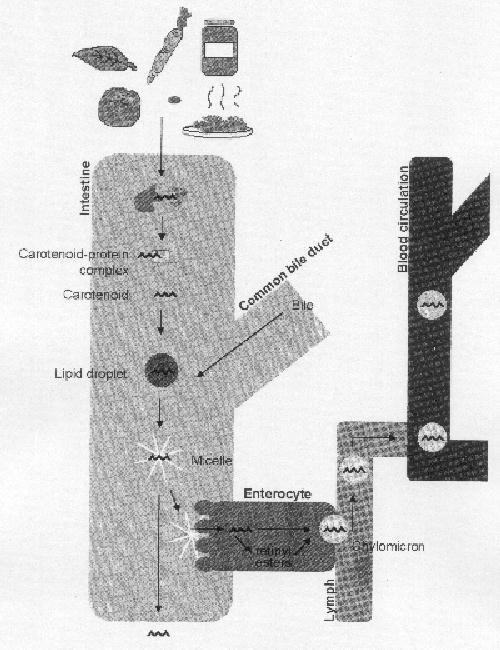
Figuur 7: Schematisch overzicht van de opname van carotenoïden (Van het Hof, 1995)
Er is onderzoek gedaan naar de opname en distributie van lycopeen in het menselijk lichaam. Lycopeen wordt op dezelfde manier verteerd als voedingsvetten. Lycopeen wordt vrijgemaakt uit de voedselstructuur en wordt in de darm opgelost in vet. Het vrijkomen van lycopeen wordt beïnvloed door de beschikbaarheid in de voedselstructuur, de deeltjesgrootte van het voedsel na het kauwen en de werking van de maag en de effectiviteit van de verteringsenzymen. Dit gebeurt in de aanwezigheid van vet en geconjugeerde galzuren (Johnson, 1998). In de dunne darm wordt lycopeen opgenomen in micellen die gevormd zijn uit voedingsvetten en galzuren (Figuur 7). Deze micellen vereenvoudigen de absorptie in de darmcellen. De absorptie van de micellen verloopt waarschijnlijk via passief transport (Stahl en Sies, 1996). In de darmcellen wordt het lycopeen opgenomen in chylomicronen. Dit zijn vesikels met een buitenste laag van fosfolipiden en een beetje apoproteïnen en een kern van vooral triglyceriden en een kleine hoeveelheid cholesterol (webpagina 6). Chylomicronen zijn verantwoordelijk voor het transport van lycopeen naar de bloedvaten via het lymfatisch weefsel. Lycopeen wordt in het bloed voornamelijk getransporteerd in LDL (low density lipoproteine) (Stahl en Sies, 1996; Johnson, 1998).
Chemische vorm
In het bloedserum en in de weefsels van de mens bestaat meer dan 50 procent van het lycopeen uit cis-isomeren (Stahl et al., 1991; Rao en Agarwal, 1999; Boileau et al., 1999; Shi en Le Maguer, 2000). Dit in tegenstelling tot voedingsmiddelen die lycopeen bevatten. Hier is 79-92 procent in de all-trans-vorm aanwezig (Boileau et al., 1999). Er is bewezen dat cis-isomeren van lycopeen beter biobeschikbaar zijn dan de trans-vorm. Dit komt waarschijnlijk omdat cis-isomeren beter oplosbaar zijn in galzuurmicellen en de voorkeur krijgen voor opname in chylomicronen (Boileau et al., 1999; Shi en Le Maguer, 2000).
In een onderzoek is gebleken dat onafhankelijk van de cis/trans-verhouding in de basisoplossing, er altijd meer dan 50 procent cis-lycopeen aanwezig is in de micellen. Dit is het gevolg van het feit dat de cis-isomeer beter oplosbaar is. Bij een basisoplossing van 100 procent trans-lycopeen vindt eerst isomerisatie plaats voordat de opname in de micellen plaatsvindt. Isomerisatie vindt echter alleen plaats in de aanwezigheid van vet. Het feit dat meer dan 50 procent van de lycopeen in de micellen in de cis-vorm aanwezig is, betekent niet dat 50 procent van alle trans-lycopeen in het voedsel deze isomerisatiereactie ondergaat. Het zou goed kunnen dat slechts een klein gedeelte van trans-lycopeen wordt omgezet en dat een groot gedeelte dus niet wordt opgenomen. Trans-lycopeen heeft een langere effectieve ketenlengte dan cis-lycopeen vanwege zijn gestrekte keten. De vorming van de cis-conformatie verkort de effectieve ketenlengte, zodat het beter oplosbaar wordt. Het is niet helemaal duidelijk waarom dit zo is. Misschien heeft dit iets te maken met de vorming van kristallijne gebieden. Cis-isomeren vormen geen kristallijne gebieden, terwijl de trans-isomeren dit wel doen. Hierdoor is het minder waarschijnlijk dat cis-isomeren aggregaten vormen en de kans wordt hierdoor groter dat ze opgenomen worden in de micellen (Boileau et al., 1999). In een tweede experiment is gebleken dat de verhouding tussen cis- en trans-lycopeen in het lichaam van ongeveer 1:1 het meest voorkomt. Deze verhouding vertegenwoordigt het natuurlijke evenwicht tussen trans- en cis-lycopeen. In de tomaat wordt dit evenwicht echter voorkomen door de celstructuur (Boileau et al., 1999). De biobeschikbaarheid van de cis-vorm is beter dan de biobeschikbaarheid van de trans-vorm. Het is bekend dat in het lichaam de trans-vorm wordt omgezet in de cis-vorm (Boileau et al., 1999). Aangezien het percentage trans-lycopeen in het voedsel veel hoger is dan het percentage trans-lycopeen in de micellen, hebben we aangenomen dat deze omzetting plaatsvindt in de darm. Het is de vraag of deze omzetting snel genoeg gebeurt. De omzetting moet namelijk plaatsvinden voordat lycopeen weer uit de darm is verdwenen. Het zou daarom beter zijn voor de opname als lycopeen in het voedsel reeds voor 50 procent in de cis-vorm aanwezig zou zijn.
De opname van lycopeen is afhankelijk van de dosis, maar dit verband is niet lineair. Wanneer een kleine hoeveelheid lycopeen, namelijk 0,35 m
mol lycopeen per kilogram lichaamsgewicht, wordt geconsumeerd, wordt 85 nmol lycopeen per liter bloed opgenomen. We hebben de oorspronkelijke absorptie op 100 procent gesteld. Wanneer de dosis 3 tot 4 keer zo groot wordt, is de absorptie van lycopeen nog slechts 50 tot 67 procent. Als de dosis hierna nog eens 2 keer zo groot wordt, is de absorptie van lycopeen nog maar 38 tot 50 procent. De toename in lycopeen in het bloed na de inname van verschillende doses lycopeen op dag 0 is weergegeven in figuur 8 (Stahl en Sies, 1992). Helaas is in dit onderzoek geen onderscheid gemaakt tussen de opname van cis- en trans-lycopeen. Een verklaring voor de verminderde toename van lycopeen bij inname van hogere doses zou kunnen zijn dat de chylomicronen reeds verzadigd zijn, zodat er geen lycopeen kan worden getransporteerd naar het bloed.
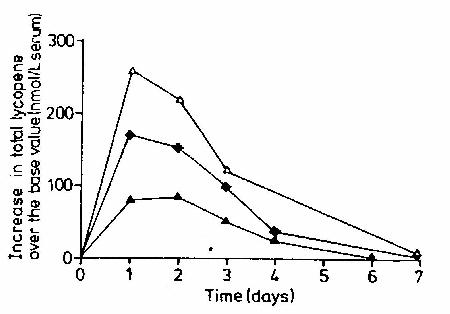
Figuur 8: De toename in lycopeenconcentraties ten opzicht van de basale waarde na de consumptie van 0,35 (dichte driehoeken
), 1,25 (¨
) en 2,5 (open driehoeken
) m
mol lycopeen per kg lichaamsgewicht (Stahl en Sies, 1992)
Ook bij herhaalde consumptie van lycopeen neemt het gehalte in het bloed steeds minder toe. De toename van het totale lycopeengehalte in het bloed na herhaalde consumptie van tomatensap met 0,35 m mol lycopeen per kilogram lichaamsgewicht is weergegeven in figuur 9 (Stahl en Sies, 1992). Als aangenomen wordt dat de toename in het totale lycopeengehalte in het bloed na herhaalde consumptie steeds minder wordt, kan worden geconcludeerd dat er een maximale waarde voor het lycopeengehalte in bloed bestaat.
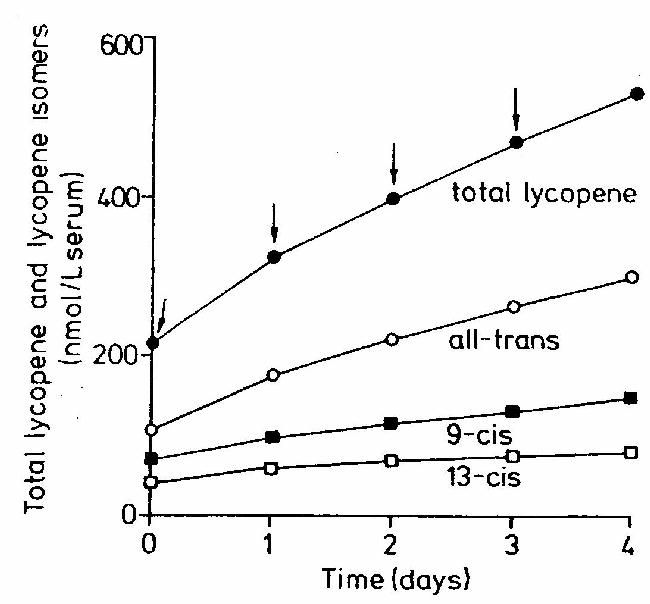
Figuur 9: Verhoging in het lycopeengehalte en zijn isomeren na herhaalde consumptie van 0,35m
mol lycopeen per kg lichaamsgewicht. De pijlen geven de tijd van consumptie aan. (Stahl en Sies, 1992) Factoren in het voedsel
Vet De biobeschikbaarheid van lycopeen uit tomatenproducten is significant hoger als de tomatenproducten worden geconsumeerd in aanwezigheid van vet (Stahl en Sies, 1996 en 1992; Porrini et al., 1998; Shi en Le Maguer, 2000). Voor alle stappen in het opnameproces van lycopeen is vet nodig, namelijk voor de vertering van het tomatenproduct, de vorming van micellen, de opname in de darm en het transport via het lymfatische systeem (Johnson, 1998). Tijdens de vertering stimuleert het vet de galvorming en zo de vorming van vetmicellen. De mate van opname van lycopeen in micellen en chylomicronen is van grote invloed op de biobeschikbaarheid. Hoe meer vet er aanwezig is, des te meer micellen en chylomicronen er kunnen worden gevormd (Porrini et al., 1998). Voor de opname van b-caroteen, een ander carotenoïde, is 5 tot 10 gram vet nodig in een maaltijd voor voldoende absorptie (Williams et al., 1998). Hierbij werd geen melding gemaakt welke hoeveelheid voldoende is. We zijn daarom uitgegaan van de aanbevolen dagelijkse hoeveelheid. Aangezien lycopeen een soortgelijke structuur heeft, nemen we aan dat deze zelfde hoeveelheid vet ook geldt voor de opname van lycopeen. b-caroteen is vitamine A. De aanbevolen hoeveelheid voor vitamine A is 0,8 tot 1 mg per dag (webpagina 7). De aanbevolen hoeveelheid voor lycopeen is ongeveer 10 mg per dag. Als dit doorgerekend wordt naar de hoeveelheid vet die nodig is voor de opname van lycopeen, zou dit betekenen dat hiervoor 50-100 gram vet per dag nodig is. Dit is wel veel. Waarschijnlijk is de relatie tussen de opname en vet dan ook niet lineair.
Polysacchariden Uit studies met proefdieren wordt gesuggereerd dat verschillende typen voedingsvezels de biobeschikbaarheid van b-caroteen verminderen. Onder deze voedingsvezels vallen onder andere hemicellulose, lignine en pectine. Eén van de mogelijke verklaringen hiervoor is dat viskeuze polysacchariden, zoals pectine, het legen van de maag vertragen en de vorming van micellen, die nodig zijn voor de absorptie, belemmeren. Hoog veresterde pectines worden vooral geassocieerd met dit effect omdat zij de viscositeit extra verhogen. Onder deze condities nemen galzouten deel aan de gelvorming van de maaginhoud en kunnen de galzouten daardoor niet meer deelnemen aan micelvorming in de darm (Rock en Swendseid, 1992). Aangezien de opname van lycopeen evenals b-caroteen plaatsvindt via micellen, wordt aangenomen dat polysacchariden de opname van lycopeen remmen. Eiwit In tomaten gaat lycopeen een interactie aan met eiwit. Dit lycopeen-eiwit-complex voorkomt de opname van lycopeen in de darm. Het verbreken van dit complex zorgt voor een hogere biobeschikbaarheid van het lycopeen (Williams et al., 1998; Shi en Le Maguer, 2000). Het precieze mechanisme hiervan is niet geheel duidelijk. In de literatuur is er weinig over te vinden. Eiwit in de dunne darm kan vetemulsies stabiliseren en vorming van micellen en hierdoor ook de opname van carotenoïden verbeteren (Castenmiller, 2000). Eiwitten kunnen uit een apolair en een polair deel bestaan. Daardoor zijn ze in staat een interactie aan te gaan met zowel polaire stoffen, zoals water, als apolaire stoffen, zoals vet. Dit zorgt ervoor dat in olie-in-wateremulsies geen fasescheiding plaatsvindt. Het eiwit bevordert de emulsievorming zodat er geen coagulatie van vetdruppels kan plaatsvinden. Kleine vetdeeltjes gaan sneller in micellen dan grote vetdeeltjes. De opname van lycopeen wordt dus versneld door eiwit. Interactie met andere carotenoïden Er wordt verondersteld dat de inname van een hoge dosis van één carotenoïde de absorptie en dus ook de biobeschikbaarheid van een ander carotenoïde tegengaat. In een onderzoek bleek dat bij de inname van drie blikjes tomatensap het b-caroteengehalte in het bloed was verdubbeld. Het lycopeengehalte in het bloed was verdrievoudigd. Het b-caroteengehalte in het sap was slechts 1/3 van het gehalte lycopeen. Dit suggereert selectieve absorptie van b-caroteen (Williams et al., 1998). Een verklaring hiervoor is dat b-caroteen en lycopeen dezelfde transportroute hebben en hiervoor competitie aangaan (Johnson, 1998). In dit onderzoek is echter niet vermeld wat het basale niveau van lycopeen en b-caroteen in het bloed is. Shi en Le Maguer schrijven dat lycopeen het meest voorkomende carotenoïde in het lichaam is (Shi en Le Maguer, 2000). Het feit dat het b-caroteengehalte in het bloed met een hoger percentage toeneemt dan het lycopeengehalte in het bloed betekent niet vanzelfsprekend dat er meer b-caroteen opgenomen wordt. We vinden de conclusie van selectieve absorptie daarom wat ver gezocht. Tegengesteld hieraan worden in een onderzoek van Van het Hof basale b-caroteengehaltes en lycopeengehaltes genoemd van respectievelijk 0,34 - 0,37 mmol / liter bloed en 0,27 - 0,28 mmol / liter bloed (Van het Hof, 1999). Deze waarden zouden betekenen dat de conclusies van het onderzoek van Williams en anderen wel correct kunnen zijn. Uit een ander onderzoek blijkt juist dat de aanwezigheid van b-caroteen de absorptie van lycopeen verbetert. Een verklaring hiervoor is dat b-caroteen de oplosbaarheid van lycopeen verhoogt (Williams et al., 1998; Johnson, 1998). Hoge inname van b-caroteen kan de absorptie van lycopeen verbeteren, maar kan ook het transport van lycopeen in LDL verhinderen (Johnson, 1998).
Cholesterolgehalte in het bloed Bij alle carotenoïden is een positief verband gevonden tussen het totale cholesterolgehalte en het gehalte carotenoïde in het bloed. Deze relatie is ook gevonden voor LDL-cholesterol. Voor HDL-cholesterol was deze relatie alleen aanwezig bij a
- en b-caroteen en lycopeen (Brady et al., 1996). Het positieve verband kan worden verklaard door het transportmechanisme van carotenoïden in het bloed. Zij worden namelijk getransporteerd door cholesterol. Het positieve verband van lycopeen met HDL-cholesterol is vreemd. Lycopeen wordt voornamelijk getransporteerd door LDL. Er is echter geen verklaring te geven voor dit positieve verband met HDL. pH van de maag In een onderzoek bij mannen en vrouwen die leden aan een ernstige vorm van atrophische gastritis bleek dat zij een zeer laag carotenoïdgehalte hadden in het bloed. Bij atrophische gastritis wordt in de maag geen of weinig zuur gevormd. Er is verondersteld dat de pH in de maag invloed heeft op de opname van carotenoïden (Tang et al., 1996). Uit een volgend onderzoek over b-caroteen bleek dat vooral het gehalte van trans-b-caroteen in het bloed bij lage pH in de maag significant hoger was dan bij een hoge pH (Tang et al., 1996) (Figuur 10).
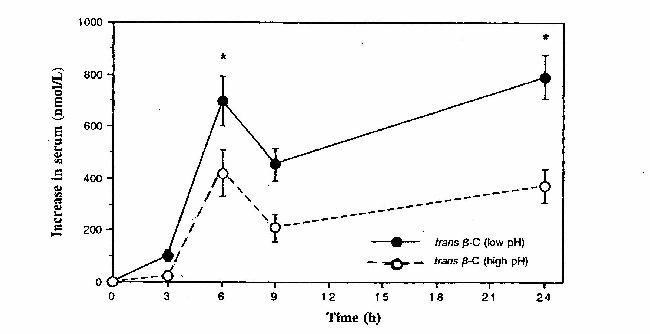 Figuur 10: Toename in serumconcentratie van trans b-caroteen bij een pH in de maag van 1,3 ±
0,1 en 6,4 ±
0,3 (Tang et al., 1996). Figuur 10: Toename in serumconcentratie van trans b-caroteen bij een pH in de maag van 1,3 ±
0,1 en 6,4 ±
0,3 (Tang et al., 1996).
Aangezien b-caroteen een soortgelijke structuur heeft als lycopeen nemen we hier aan dat het mechanisme van opname hetzelfde is. Er worden meerdere verklaringen gegeven voor het hogere gehalte in het bloed bij lage pH. Ten eerste wordt een hoge pH in de maag geassocieerd met kolonisatie van bacteriën in het maagdarmkanaal. Er is echter geen bewijs gevonden dat bacteriën de opname van b-caroteen zouden kunnen tegengaan. De meest waarschijnlijke verklaring is dat een verhoging van de concentratie H+ de negatieve lading aan het oppervlak van de micellen en van het membraan van de darmcellen onderdrukt en hierdoor de diffusie van de micellen vereenvoudigt. Normaal gesproken zorgt leging van de maag in de darm voor een pH-verlaging in de darm. Wanneer de pH in de maag echter hoger is, dan is de daling van de pH in de darm kleiner. Vanwege deze verminderde daling worden de negatieve ladingen aan het oppervlak van de micellen hoger, zodat de diffusie vertraagd wordt. Vertraging van de opname hoeft echter niet te betekenen dat b-caroteen slechter wordt opgenomen. Het uiteindelijke verschil in opname tussen lage en hoge pH wordt uit dit onderzoek niet geheel duidelijk (Tang et al., 1996).
Geslacht Er zijn geen verschillen gevonden in de lycopeengehaltes in het bloed tussen mannen en vrouwen. De lycopeengehaltes in het bloed waren bij mannen en vrouwen positief gecorreleerd met de inname in het voedsel. Dit suggereert dat er geen verschillen zijn in de absorptie of afbraak van lycopeen tussen de geslachten (Johnson, 1998). Bij vrouwen is er echter wel variatie in het lycopeengehalte in het bloed tijdens de verschillende fases in de menstruatiecyclus. De menstruatiecyclus bestaat uit twee fases. In de eerste fase, de folliculaire fase, vindt groei van de follikels plaats. Uiteindelijk wordt er één follikel rijp. Die follikel wordt een graafse follikel genoemd. Aan het eind van deze fase, op dag 14 van de menstruatiecyclus, barst de graafse follikel open en komt het eitje vrij. Dit wordt ovulatie genoemd. In de luteale fase onstaat uit deze lege follikel het gele lichaam. Dit lichaam produceert hormonen, waardoor er geen nieuwe follikels kunnen rijpen. Bovendien zorgt dit lichaam voor de innesteling van het eitje in de baarmoederwand. Aan het eind van de luteale fase, dag 28 van de menstruatiecyclus, wordt het gele lichaam afgebroken en begint de menstruatie. Tijdens de menstruatie wordt het opgebouwde gedeelte van de baarmoeder weer afgestoten. Op dag 1 van de menstruatie begint de nieuwe folliculaire fase (Fox, 1996). Het totale en het LDL-cholesterol is het hoogste tijdens de vroege folliculaire fase, terwijl het HDL-cholesterol piekt tijdens de late folliculaire fase. Doordat lycopeen vooral door LDL wordt getransporteerd, is de concentratie het hoogst in de luteale fase, dus na de piek van het LDL (Tabel 7) (Forman et al., 1996). Tabel 7: Concentraties lycopeen en cholesterol in mmol/liter tijdens de verschillende fasen van de menstruatiecyclus
| |
Menstruatie |
Vroege folliculaire fase |
Late folliculaire fase |
Luteale fase |
|
Lycopeen |
1,023 ±
0,034 |
1,060 ±
0,019 |
1,023 ±
0,022 |
1,135 ±
0,022 |
|
Totaal cholesterol |
4,34 ±
0,07 |
4,38 ±
0,03 |
4,36 ±
0,04 |
4,28 ±
0,04 |
|
LDL-cholesterol |
2,61 ±
0,05 |
2,70 ±
0,04 |
2,61 ±
0,04 |
2,58 ±
0,04 |
|
HDL-cholesterol |
1,41 ±
0,02 |
1,42 ±
0,01 |
1,50 ±
0,01 |
1,45 ±
0,02 |
Het is vreemd dat er zo'n lange tijd zit tussen de LDL-piek en de lycopeenpiek. De verwachting is namelijk dat direct na de LDL-piek er meer transport van lycopeen plaatsvindt, waardoor het gehalte in het bloed enorm toeneemt. Het is eigenaardig dat dit transport meer dan een week zou duren. Het zou kunnen dat het lycopeen gedurende deze tijd wordt opgeslagen in de lymfen. Hier is echter niets over bekend. Leeftijd Als de leeftijd toeneemt, neemt bij de meeste carotenoïden het gehalte in het bloed toe. Bij lycopeen is dit echter omgekeerd. Het lycopeengehalte in het bloed neemt af bij toenemende leeftijd. Een verklaring hiervoor is dat jongeren meer lycopeen-rijke voedingsmiddelen zoals pizza en ketchup consumeren. Het dieet verklaart echter slechts een deel van dit omgekeerd evenredige verband (Johnson, 1998). Misschien wordt de opname van lycopeen slechter bij toenemende leeftijd. Gedragsfactoren
Roken Sigarettenrook produceert vrije radicalen en kan oxidatieve stress veroorzaken. De verwachting is dat bij rokers het lycopeengehalte in het bloed hierdoor lager zou zijn. Het lycopeengehalte in het bloed van rokers is echter niet verschillend van het lycopeengehalte in het bloed van niet-rokers. Een verklaring hiervoor zou zijn dat rokers meer lycopeen consumeren. Dit is echter onwaarschijnlijk (Johnson, 1998). Er is onderzoek gedaan naar het lycopeengehalte in het bloed direct voor en direct na het roken van een sigaret. Hieruit is gebleken dat het lycopeengehalte direct na het roken van een sigaret veertig procent is gedaald vergeleken met het lycopeengehalte direct voor het roken van een sigaret (Rao en Agarwal, 1998). Uit deze twee onderzoeken is niet duidelijk gebleken of het roken van sigaretten invloed heeft of niet. Het blijkt dat het lycopeengehalte direct na het roken is gedaald, echter over het algemeen zijn de lycopeengehaltes van rokers en niet-rokers gelijk aan elkaar. Het lijkt erop dat het lichaam de daling van het lycopeengehalte direct weer compenseert, het is echter niet duidelijk hoe. Alcoholgebruik Evenals sigarettenrook kan de consumptie van alcohol een bron van oxidatieve stress zijn. Daarom kan de concentratie van antioxidanten, zoals lycopeen, verlaagd zijn in de aanwezigheid van alcohol. De resultaten die hier over zijn gevonden zijn echter niet consistent. Het lycopeengehalte lijkt minder te worden beïnvloed door de consumptie van alcohol dan de andere carotenoïden (Johnson, 1998). Medicijngebruik Er zijn verschillende medicijnen die een invloed kunnen hebben op de opname van lycopeen. Voorbeelden hiervan zijn cholesterolverlagende medicijnen en medicijnen die de pH van de maag beïnvloeden. Cholesterolverlagende medicijnen zorgen ervoor dat het LDL-cholesterolgehalte verlaagd wordt. Het LDL zorgt vooral voor het transport van lycopeen in het bloed. Bij verlaging van het LDL-gehalte wordt het transport verminderd en zo ook het lycopeengehalte in het bloed (Rao en Agarwal, 1999). Omeprazole is een medicijn dat tijdelijk de maagzuursecretie kan verhogen (Tang et al., 1996). Dit medicijn verlaagt dus de pH in de maag en zou dus de opname van lycopeen kunnen versnellen. Gebruik van voedingssupplementen Supplementen die b-caroteen of andere carotenoïden bevatten, kunnen de opname van lycopeen beïnvloeden (Brady et al., 1996). Uit eerdere onderzoeken zijn hierover echter tegenstrijdige resultaten gevonden. Het is dus ook niet duidelijk wat de precieze werking van voedingssupplementen op de opname van lycopeen is. Vezelrijke voedingssupplementen remmen waarschijnlijk de opname van lycopeen. Hierbij wordt aangenomen dat vezels een soortgelijke werking hebben als pectine.
Het blijkt dus dat er veel factoren zijn die invloed hebben op de biobeschikbaarheid van lycopeen. Factoren zoals de chemische samenstelling, de dosis en de aanwezigheid van vet in het voedsel hebben relatief veel invloed, terwijl factoren zoals pH van de maag en het geslacht of de leeftijd van de consument nauwelijks invloed hebben. Bij het vergroten van de biobeschikbaarheid van lycopeen in tomatenproducten is het van belang om vooral rekening te houden met deze belangrijkste factoren.
|

| |
|































